
早速続きに参りましょう。
前回は、CRISPR-Cas9(クリスパー・キャスナイン)という最先端技術の主要登場人物の1つ、Cas9というタンパク質についてちょろっと見ていました。
こいつは何かというと、詰まる所、「DNAを切断する能力をもった生体分子マシーン」だということですね。
とはいえDNAを切断するマシーンというのは、実は生体内には他にもウジャウジャいるんですけど、一体こいつの何がスゴいのか…?
それがズバリ、「自分の好きな場所のみを、正確に切断することができる」という、切る対象が特定のDNA配列のみに限られているわけではない(だけど、ランダムにメチャクチャ切りまくるのではなく、決まった場所のみを切れる)という自由度に他ならないという話でした。
もちろん現実的には「どこでも必ず切れる」というわけではないんですけど、基本的にはほぼ無限大の自由度で、好きな遺伝子を自由自在に切ることが可能となっている(その選択の余地がある)ということですね。
…なぜそんなことが可能になるのか…?
改めて、前回も貼らせていただいた、Thermo Fisherの解説記事の図に一部補足を加えたものを貼らせていただき、順に説明していきましょう。
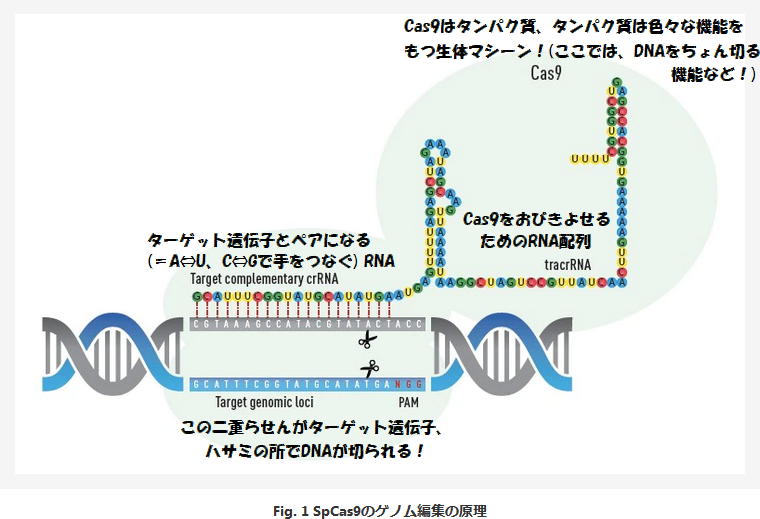
まず前回も触れた通り、Cas9は薄い緑で描かれた丸いやつですけど、こいつはDNAを切断するハサミ能力の他に、「クリスパーRNAという特別な配列をもったRNAとくっつく」という能力ももっているのです。
そのクリスパーRNAというのが、図にあるA, C, G, Uの丸いボール(A青・C赤・G緑・U黄)がつながったものになりますが(厳密にはこの1つながりのRNA全体は (前回も書いていた通り) sgRNAと呼ばれることが多いですけど、名前はマジでどうでもいいので、クリスパーに登場するRNAということで、分かりやすくクリスパーRNAと書いちゃいましょう。やっぱり、細かい名前とかはまず気にせずに大まかなコンセプトを掴んで、正確な用語などは元記事やその他の解説記事で学ぶのがいいのではないかな、と思います。)……
…と相変わらず余談の括弧書きが長くなったので仕切り直すと、この約100塩基のRNA(この図だと、104塩基ですね)の内、具体的にCas9とくっつくというか、Cas9をおびきよせる役割をもった部分が、右半分のtracrRNA(トレーサーRNA)という配列になります。
改めて、名前はまぁどうでもいいので、呼び名はともかく機能についてを強調したい限りですけど、このクリスパーRNAは大まかに2つの領域からできていて、後ろ半分が「おびきよせ配列」であるという理解でOKだということですね。
(なお、以前の記事でも何度か触れていた通り、DNAやRNAには向きがあるので、普通は左から右に進むように描かれますし、実際この図のRNAもそうなっていますが、こいつはGで始まり最後UUUUで終わる一つながりのRNA分子だということです。)
…と、「おびきよせ配列」とかいっても、そういえば僕なら事前知識がない場合、初見で多分こういうことを思っただろうな……って点に触れてみると、
「『おびきよせる』って何だよ。このRNAとやらは、怪電波でも発してるのか?」
…みたいな疑問、まぁ僕だけかもしれませんけどもしかしたら人によっては感じるのではないかと思うのですが、これは言うまでもなくそんなわけはありませんでして……
実際の細胞内や試験管内で、分子というものは溶液内を(熱運動で)飛び回りまくっており、たまたま偶然Cas9とトレーサーRNAとが接触したら(近付いたら)、まるで磁石のN極とS極のように、両者はくっついて離れない性質をもつ、という形なわけですね。
ちょうど、以前この辺の記事でも似たようなことを語っていたんですけど、割とよく書けていたので、せっかくなので自分の記事から該当部分を引用しておくとしましょう。
このときは、T7 RNAポリメラーゼという、DNAと結合するタンパク質の話ですが、似たようなものですね。
…ちなみに前回は関連して、パックマンっぽいRNAポリメラーゼがリング状のプラスミドにくっついてる図を適当に作って載せていましたが、これも改めて、ああいう図を見ると僕なんかは度々「いやだから、『プロモーターを発見!』とか、なんでそんなことが可能なんだよ!?分子に目がついてるわけでもないんだろ?!そんな見てきたかのようにいわれても、にわかには信じがたいぜぇ~!!」と、まぁそんなに毎度必ずつっかかってたわけでもないですけど、やっぱり不思議に思うわけです。
一応これについてはもう何度か書いている話ですけど、結局RNAポリメラーゼもプラスミドも細胞内では熱運動をしているというか、適当に細胞内液を漂っている物質であって、それぞれ結構な数がありますから、たまたまぶつかり合うこともあるわけですね。
で、たまたま接触したときに、例えばT7 RNAポリメラーゼなら、T7プロモーター(=TAATACGACTCACTATAというDNAの配列)とかなり強く結合する性質をもっているので(※)、T7 RNAポリメラーゼとT7プロモーターが偶然たまたま近付いたその瞬間に上手いことガッチリとくっつきあって離れず、そこから転写が始まる、ってメカニズムになっているという話なのです。
(※注)まぁそれも、「なんでそんな都合よく強く結合できるんだよ?」って疑問にはなるわけですけど、化学的にというか物理的にというか、両者の構造に起因する分子間相互作用とか、もっと細かく表面電荷の分布による何やらとかいった小難しい話で、こいつらはペアを作りやすい組み合わせになってるんです、としかいえない話かもしれませんね。
磁石のN極とS極はくっつっきますが、まぁその仕組みは磁力線うんぬんの小難しい話で説明が可能であっても、究極的には「なぜくっつくのか?」は「知らん。この世はそうなっとんねん、としかいえん話よ。とにかく実際にくっつくんだから、仕組みはともかく、受け入れるしかあるめぇ」って話ともいえる感じでしょうか。
…そんなわけで、Cas9は、クリスパーRNAの右半分(後ろ半分)の力によって、このRNA分子のそばに引き寄せられて、ちょうど図のような位置関係に配置されることになるというわけなのです。
(くっつく場所も決まっているということですね。絵にすると、ちょうど上のような図みたいになっている形です。)
そしてそのおかげで!
見事に、CRISPR RNAとCas9との共同作業によって、「好きな配列を切ることのできる仕組み」が爆誕する所へと、話がつながっていくのです…!!
…と、実に中途半端な所で恐縮ですが、「記事は短いほど良い」「1記事で1つ新しいことに触れるぐらいでちょうどいいんじゃない?」という観点から、この続き・左半分のいわば「認識配列」については、また次回にまわさせていただくといたしましょう。
って本当はちょっと最後まで書く時間がなかっただけなんですけど(笑)、むしろこのぐらいの方がマジで1記事としてちょうどいい分量かもしれませんね。
というわけで、次回はこのクリキャスシステムを神がかり的なものにしている最大の功労者・RNAの前半分について見ていこうと思います。